|
| ||||||
|
| |||||
|
Тема 3. ОТДЕЛ ПЛАУНОВИДНЫЕ (LYCOPODIOPHYTA, ИЛИ LYCOPHYTA) Общая характеристика Плауновидных. 2. Классификация Плауновидных. Класс плауновые (Lycopodiopsida). Класс полушниковые (Isoetopsida). 3. Циклы развития Плауновидных.
Общая характеристика Плауновидных Плауновидные представляют собой самую древнюю группу из числа ныне живущих высших растений. Все плауновидные имеют олиственные побеги и весьма своеобразные корни. Наиболее характерной особенностью плауновидных является микрофилия, под которой понимаются относительно мелкие размеры листьев, анатомо-морфологическая простота и их энационное происхождение. Листья плауновидных имеют цельную листовую пластинку и несколько расширенное основание, иногда в виде подушечки. У некоторых плауновидных на расширенном основании листа формируются выросты в виде язычка (лигулы). Первые листья молодого растения не имеют жилок. В ходе онтогенеза лист развивается из одной поверхностной клетки стебля. Все это указывает на то, что в ходе филогенеза листья плауновидных возникли как поверхностные выросты, или энации, на осевых органах, что делает их среди прочих высших растений уникальными. Некоторые авторы энационные листья плауновидных называют филлоидами, т.е. трактуют их как листоподобные органы. Для плауновидных, как и для всех прочих споровых растений, характерно отсутствие главного корня; все корни занимают на побегах боковое положение. Уникальность плауновидных заключается в том, что и побеги, и корни обладают самым древним - дихотомическим типом ветвления. Спорангии плауновидных формируются на верхней стороне спорофиллов, которые у подавляющего числа видов образуют специализированные спороносные побеги - стробилы в виде колосков. Плауновидные объединяют как равноспоровые, так и разноспоровые растения.
Классификация Плауновидных Классификация плауновидных испытывает сложности из-за наличия переходных форм, особенно среди вымерших видов. Внутри отдела выделяются 2 класса - плауновидные (Lycopodiopsida) равноспоровые и полушниковые (Jsoetopsida), в основном разноспоровые.
Класс Плауновые (Lycopodiopsida) Этот класс объединяет равноспоровые растения, не имеющие в основании листа подушечек. Он включает порядки астероксиловых (Asteroxylales), дрепанофикусовых (Drepanophycales) и плауновых (Lycopodiales). Два первых порядка представлены исключительно вымершими формами. Из числа вымерших плауновидных один из наиболее древних - род астероксилон (Asteroxylon), известный из среднего девона. Судя по сохранившимся надземным частям, это были невысокие травянистые растения, обладавшие неравной дихотомией. Как главный, так и боковые побеги были обильно покрыты мелкими шиповатыми листьями. Внешне эти растения напоминали современные плауны. К каждому листу подходила жилка, которая оканчивалась в самом его основании, не входя в листовую пластинку, т.е. по происхождению эти листья были энациями. Между листьями на коротких ножках беспорядочно располагались спорангии почковидной формы, которые вскрывались на вершине продольной щелью, т.е. спорангии еще не были связаны непосредственно с листьями. Стебель с поверхности был покрыт типичной эпидермой с устьицами. Большую часть стебля занимала кора, которая подразделялась на периферическую, состоявшую из плотных паренхимных клеток, и внутреннюю, с хорошо выраженной аэренхимой. В центре располагалась актиностела; на поперечном срезе она имела вид расходящихся от центра лучей экзархной ксилемы и участков флоэмы между ними. (Своим названием этот род обязан звездчатой форме ксилемы.) Подземные органы астероксилона неизвестны. Своеобразие плаунов заключается в их исключительной примитивности как спорофитов, так и гаметофитов, что связано с большой древностью этого рода. Это самый древний род из всех высших растений на Земле. К числу примитивных признаков относятся: 1) дихотомическое ветвление стеблей и корней; 2) отсутствие строгой детерминированности при заложении побегов и корней; 3) отсутствие придаточных корней; 4) анатомическое сходство в строении стеблей и корней; 5) наличие самых примитивных типов стели у ряда видов - протостелы и актиностелы; 6) большая продолжительность жизни гаметофитов; 7) наличие длинной шейки архегония. Все это позволяет рассматривать род плаун как истинно живое ископаемое растение.
Класс полушниковые (Isoetopsida)
Данный класс распадается на 4 порядка: Селягинелловые (Selaginellales), Протолепидодендровые (Protolepidodendrales), Лепидодендровые (Lepidodendrales), Полушниковые (Isoetales). Причем современные виды представлены только среди селягинелловых и полушниковых. К первому порядку относится всего 1 род селягинелла (Selaginella), насчитывающий около 700 видов. Порядок полушниковых включает 2 современных рода - стилитес (Stylites), представленный одним видом и полушник (Isoetes), насчитывающий около 70 видов.
Циклы развития Плауновидных
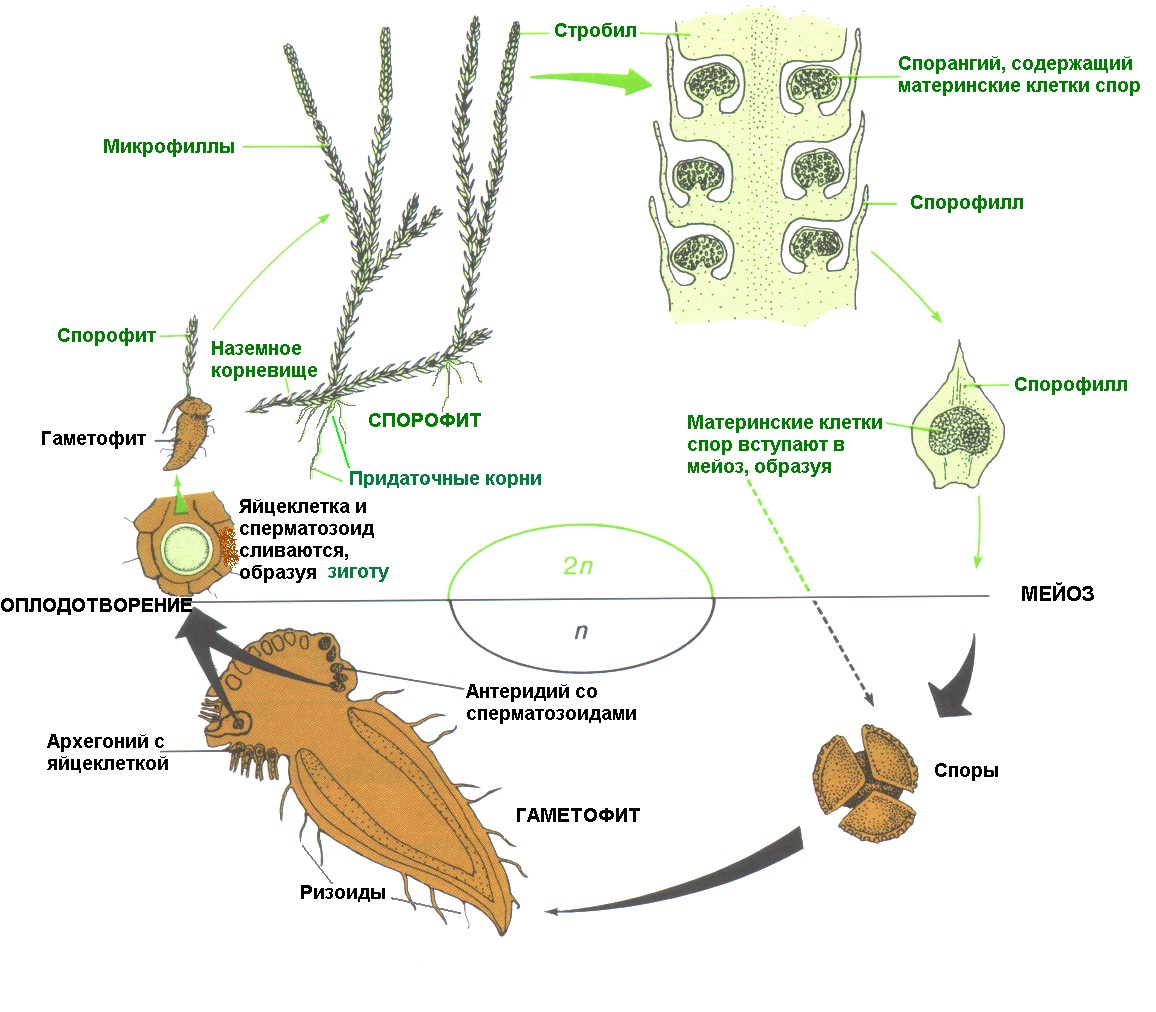
Цикл развития плауна (рис.5). Бесполое размножение плаунов осуществляется с помощью спор, образующихся в спорангиях почковидной формы. В типичных случаях спорангии, расположенные на короткой ножке, развиваются на верхней стороне спорофилла. У большинства плаунов на вершине особых побегов располагаются стробилы в виде колосков, длина которых у разных видов варьируют от 0,5 до 40 см. В спорангиях в результате редукционного деления образуются мелкие тетраспоры, содержащие масло и хлоропласты. Споры прорастают через 3-8 лет после высыпания из спорангия. Из споры возникает новое поколение - гаплоидный гаметофит, или заросток, на котором формируются половые органы. Заросток по-латыни называется проталлиум. На них образуются антеридии и архегонии. Существует два типа прорастания спор - подземное и надземное. В почву спора попадает либо с токами воды, либо прикрывается растительным опадом. Рост и развитие подземного гаметофита, размеры которого измеряются миллиметрами, возможен только при условии внедрения в него гиф гриба, за счет которого он питается. Развитие гаметофита происходит чрезвычайно медленно, созревание архегониев и антеридиев завершается через 6-15 лет после начала прорастания споры. При надземном прорастании развитие гаметофита завершается в течение нескольких дней. По форме гаметофиты могут быть цилиндрическими, червеобразными, редьковидными, похожими на волчок. У одних видов, например у баранца, гаметофит лишь частично скрыт в земле, при этом его надземная зеленая часть способна ассимилировать, а подземная бесцветная - обеспечивает водоснабжение. У других видов бесцветный гаметофит полностью находится в земле, но при извлечении на свет он зеленеет. У большинства же видов бесцветные подземные заростки на свету не зеленеют, т.е. полностью перешли к микотрофному типу питания. Гаметофиты у плаунов обоеполы; архегоний и антеридии располагаются на их верхней стороне. Первыми появляются антеридии, полностью погруженные в ткань гаметофита. Сперматозоиды плаунов самые мелкие среди всех высших растений. Они имеют червеобразную форму и снабжены парой длинных жгутиков. Архегоний также погруженные, над поверхностью заростка выступают лишь клетки шейки. После оплодотворения зигота делится на 2 клетки. Верхняя из них вытягивается и превращается в подвесок, который вдавливает развивающийся зародыш в ткань гаметофита, обеспечивая ему питание. Нижняя клетка при делении образует сначала мощную гаусторию, а затем формирует зародыш. Таким образом, на первых этапах развития зародыш молодого спорофита, так же как у мохообразных, питается за счет гаметофита. В отличие от них у плаунов очень рано формируются основные вегетативные органы - побег и корень, которые обеспечивают дальнейшее независимое существование спорофита.
Рис.5. Жизненный цикл плауна
Цикл развития селягинеллы (рис.6). Бесполое размножение у селягинелл осуществляется спорами разной величины - макро- (или мега-) и микроспорами. Микроспоры в большом числе возникают в микроспорангии, расположенном на микроспорофилле, а мегаспоры в числе 1-4 формируются в мегаспорангии несколько больших размеров, которые возникают на мегаспорофиллах. В большинстве случаев микро- и мегаспорофиллы объединяются в одном стробиле в форме колоска, у других - имеются лишь спороносные зоны, чередующиеся с участками вегетативных побегов. Прорастание микроспор начинается внутри микроспорангия. После первого деления образуются две неравноценные клетки - одна маленькая, представляющая собой остаток вегетативного тела заростка и поэтому называемая проталлиальной. Вторая клетка, большая, дает начало антеридию и поэтому называется антеридиальной. Антеридий состоит из восьмиклеточ-ной стенки и 2 или 4 сперматогенных клеток, формирующих сперматозоиды. Такой микроскопический мужской гаметофит, состоящий из 11 или 13 клеток и покрытый оболочкой микроспоры, вылетает из раскрывшегося микроспорангия. Подобно пылинке семенных растений гаметофит разносится ветром, поэтому его тоже можно назвать пылинкой. Деление мегаспоры также начинается под защитой ее оболочки, но в отличие от мужского заростка женский заросток состоит из большого числа проталлиальных клеток. Под бороздой споры деление клеток происходит более интенсивно, поэтому в этом месте оболочка мегаспоры лопается, и гаметофит выступает за ее пределы. Здесь формируются несколько погруженных в ткань гаметофитов архегониев. Близ каждого архегония возникает небольшой бугорок с пучком длинных ризоидов. У большинства видов селягинеллы женский заросток выпадает из спорангия и попадает на почву. В этом случае ризоиды выполняют функцию поглощения воды, необходимой для оплодотворения. Сперматогенные клетки, многократно делясь, образуют от 128 до 256 двужгутиковых сперматозоидов, выплывающих через лопнувшую оболочку микроспоры. Оплодотворение яйцеклеток и развитие зародыша происходят на материнском растении под защитой стенки мегаспорангия. В биологическом смысле этот процесс напоминает формирование семян у семенных растений, хотя и не является его гомологом. У селягинелловых, как и во всех прочих группах разноспоровых растений, прослеживается закономерная связь между разноспоровостью, раздельнополостью гаметофитов и их значительной редукцией, которая у семенных растений достигла наибольшего выражения. Следовательно, можно ожидать, что все эти три процесса имеют важное биологическое значение. При разноспоровости в микроспорангиях образуется большое количество очень мелких спор, дающих начало сильно редуцированным мужским гаметофитам - пылинкам, которые легко разносятся ветром. Массовость гаметофитов повышает эффективность полового процесса. Большая скорость развития мужских гаметофитов и значительная их редукция обеспечивают им защиту от высыхания благодаря кутинизированной оболочке микроспоры. Кроме того, незначительные размеры гаметофита способствуют экономии пластического материала. В мегаспорангий происходит резкое сокращение количества мегаспор до 4-1 в сочетании с увеличением их размеров и содержанием в них питательных веществ. Это способствует развитию более крупного, более жизнеспособного женского гаметофита, который также находится под оболочкой мегаспоры. После оплодотворения яйцеклетки на этом гаметофите может сформироваться полноценный зародыш нового спорофита. Раздельнополость гаметофитов обеспечивает новому поколению расширение, обогащение генетического потенциала.
Рис.6. Жизненный цикл селагинеллы
| ||||||
|
| ||||||
| Сайт создан по технологии «Конструктор сайтов e-Publish» | ||||||